
(Animated Artificial Life)
3.4 Sexual Swimmers
In the latest series of simulations, physics is shifted from a terrestrial model to an aquatic one. Dimensionality is brought down from three to two, but the model is deepened in another respect: reproduction is spontaneous. To ground the simulation more in a physical representation, the concept of a "generation" is removed. Instead of updating the entire population in discrete stages (a schedule determined by a "creator"), this model allows reproduction to occur asynchronously and locally among creatures able to reach proximity to each other in a spatial domain. The notion of an explicit fitness function for selection is removed [Ventrella, 1996].
Whereas the previous simulations explicitly rewarded creatures for speedy locomotion, this simulation lets them reward themselves by mating. Speedy locomotion therefore becomes a prerequisite (along with many other behaviors), and not the sole objective.
3.4.1. The Energy Cycle
Energy exists in the fluid medium in ambient form, and is periodically converted into bits of food distributed randomly in the fluid. These food bits can be eaten by the swimming creatures to attain energy. They use this energy to move body parts and exert force against the fluid medium. Their energy levels decrease in proportion to physical exertion. Creatures are by default interested in mating, and pursue other creatures for this purpose. However, if a creature's energy dips below a specific threshold, it changes its mental state and pursues a nearby food bit instead of a mate. Swimming creatures who are genetically disposed to erratic motion waste large amounts of energy, and thus spend more of their lives pursuing food. They have less time to dedicate to propagating. Abnormally erratic creatures are usually not even able to reach a food source before losing all their energy, and so they usually die off early-on in evolution. Over evolutionary time, the most energy-efficient swimmers survive. It should come as no surprise that these swimmers, having evolved for energy-efficiency, are also beautiful to watch.
3.4.2. Physics and Genetics are Linked
It is important to point out that in this simulation, genetic evolution is totally reliant on the physical model which animates the creatures. Not only does it take Two to Tango, but they have to be able to swim to each other as well (more specifically, at least one has to be able to swim to the other). In this simulation, genotypes are housed in physically-based phenotypes, which, over evolutionary time, become more efficient at transporting their genotypes to other phenotypes, thereby enabling them to mate and reproduce their genes. This is the selfish gene at work. The emergence of locomotive skill becomes meaningful therefore in the context of the local environment: it becomes physically grounded, and the fitness landscape becomes dynamic.
Locomotion behavior evolves by way of local matings between creatures with higher (implicit) fitness than others in the local area. Often a maturing population will separate into two distinct regions in phenotype space, before it is taken over by one phenotype, corresponding to different areas of the fluid medium, where genotypes cannot readily mix.
Figures 10a and b illustrate two swimming styles which emerged from a simulation.

Figure 10. Sequences of images of swimming strategies: (a) paddling-style, and (b) undulating-style, which emerged through evolution
3.4.3. Turn That Body
A swimming creature spends most of its life pursuing a potential mate. That potential mate may possibly be pursuing its own potential mate, and so it is often a moving target. A swimming creatures has to be able to continually orient itself in the direction it wants to go. A reactive system was designed which allowed a creature to detect how much it would need to turn its body while swimming. Essentially, the creature senses the angle between the direction of its goal and its own orientation as the stimulus to which it reacts, as shown in Figure 11. This angular relation to the goal is dynamic, constantly changing as pursuer and pursued move throughout the fluid.
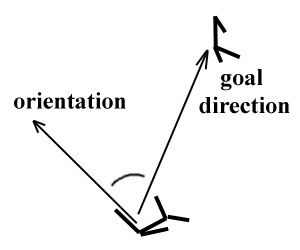
Figure 11. Turning Stimulus
As opposed to Walker, which experiences a "magic pull" in the direction of its goal, these creatures must use this sense to affect their own autonomous means of propulsion, using friction with the surrounding fluid.
Since the number of possible morphologies and motion strategies in these creatures is large, it would be inappropriate to top-down-design a turning mechanism. Since turning in the plane for a 2D articulated body can be accomplished by modulating the phases and amplitudes of certain part motions, it was decided that evolution should be the designer, since evolution is already the designer of morphology and motion. Thus, modulator genes (such as those listed in section 2.3.7) were implemented which can affect the amplitudes and phases in each part's motions in response to the stimulus.
A commercial product was derived from this simulation called "Darwin Pond" [RSG, 1997]. It enables users to observe hundreds of swimming creatures, representing a large phenotypic space of anatomies and motions. They can be observed in the Pond with the aid of a virtual microscope, allowing panning across, zooming up close to groups of creatures, or zooming out to see the whole affair. One can create new creatures, kill them, move them around the Pond by dragging them, inquire their mental states, feed them, remove food, and tweak individual genes while seeing the animated results of genetic engineering in real-time. Figure 12 shows a screen shot of the basic Darwin Pond interface.
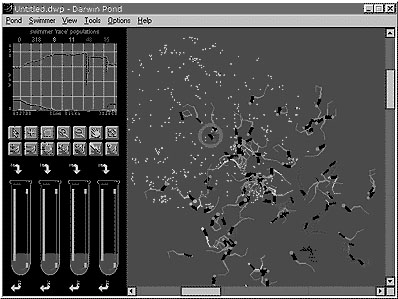
Figure 12. Darwin pond interface
A variation on this simulation, called "Gene Pool" was developed, and included a larger genotype-phenotype scheme, variable thickness in body parts, a deeper physics, and a conserved energy model. Figure 13 illustrates a collection of these 2D swimming creatures (before evolution has had any effect of body plan). They are called "swimbots."

Figure 13. Swimbot anatomies
2.4.4. What's Sex Got To Do With It?
Gene Pool deepens the simulation further by introducing mate choice in the process of reproduction. In this simulation, not only is the evolution of form and motion more physically grounded, and subject to the local conditions of the simulated world, but the aesthetic criteria which determine the "sexiest" motions also become a factor for reproduction.
What does "sexy" mean? Sexual selection in evolution is responsible for phenomena such as the magnificent peacock tail, and the elaborate colorful dances of many fish species, who sport curious body parts, adapted for mate attraction. Attraction-based features in many species may be so exaggerated and seemingly awkward that one would assume they decrease the overall fitness of the species, in which locomotion efficiency, foraging, and security are important. But the mere existence of these remarkable sexual features is proof that they are not fatal to the survival of the species. An interesting discussion on the topic of sexual selection and the biology of beauty can be found in [Moller, 1997].
Computer models developed by Todd and Miller [1993], in which sexual selection drives a population of simulated organisms into arbitrary regions of phenotype space, supplied motivation to explore this intriguing side of evolution. A question was posed: could mate preferences for arbitrary phenotypic features inhibit the evolution of energy-efficiency in a population of locomotive creatures? For instance, if all creatures in the population were attracted to slow-moving bodies, and only mated with the most stationary creatures in the population, would locomotion skill still emerge? If so, what kind? What if they were all attracted to short, compact bodies? Would the body plan become reduced to a limbless form, incapable of articulated locomotion? If they were attracted to wild, energetic motion, would the resulting swimbots be burning off so much energy that most die before reproducing?
As an experiment, a simulation was built, including a variety of mate preference criteria determining which swimbots would be chosen [Ventrella, 1998]. At the point of choosing a potential mate, a swimbot takes "snapshots" of each swimbot within its local view horizon, and then compares these snapshots and ranks them by attractiveness. It chooses the most attractive swimbot as its goal and then begins to pursue it. Criteria settings include:
* attraction to long bodies (length is measured as the greatest distance between any two body parts)
* attraction to lots of motion (motion is measured as the sum of all instantaneous speeds of all body parts)
* attraction to bodies which are "open" (openness is measured as the sum of all distances between parts)
* attraction to massive bodies (massiveness is measured as the sum of all body part areas)
The inverses of each of these phenotypic characteristics are also tested, making a total of eight attractiveness criteria. As expected, in simulations in which length was considered attractive, populations with elongated bodies with few branching parts emerged. In these populations, locomotion was accomplished by a tiny paddle-like fin in the back of the body, or by gently undulating movements. Figure 14 illustrates a local cluster of swimbots resulting from this simulation.

Figure 14. "Long is Beautiful"
Attraction to open bodies resulted in creatures which spent a large part of their periodic swimming cycles in an "open" attitude, as shown in Figure 15. In this illustration, 13 images are shown in Muybridge form to illustrate approximately one swim cycle, sequenced from left-to-right. The creature is swimming in the direction towards its lower right. Notice that the stroke recovery (the beginning and end of the sequence) demonstrates an exaggerated "opening up" of the body. Having observed hundreds of swimming cycles in evolved populations, I can easily say that this is uncommon. It is likely that this behavior emerged as a by-product of the attractiveness criteria for the following reason: at the moment a creature is ready to take snapshots to size up potential mates within its view, the potential mates are all in various arbitrary stages of their swim cycles. So it is advantageous to be attractive as much as possible throughout the entire swim cycle. The more open a swimmer's body is, and the longer amount of time it is open throughout the swim cycle, the more attractive on average it will be.
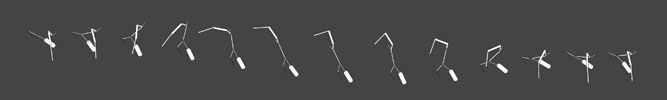
Fig. 15. Swimbots which evolved through mate preferences for open bodies.
These experiments show how mate preference can affect the evolution of a body plan as well as locomotion style. The effects on energy-efficiency seem to be non-trivial: locomotion strategies emerge which sometimes take advantage of attraction criteria yet are still efficient in terms of locomotion. As usual with most artificial life simulations, the phenotype space has unexpected regions which are readily exploited as the population adapts(regions which a top-down designer might not think to visit.
3.4.5. Just Sit and Look Pretty
To be "fit" in this world doesn't necessarily mean to be fast, good at turning, or energy-efficient. It also includes being attractive. In fact, using some attractiveness criteria (such as lack of movement, or short bodies), it was found that populations could easily evolve to consist mostly of immobile creatures with hardly any anatomy. In such evolved populations, there are a few efficient swimmers who appear to be responsible for most of the mating. These individuals seem to be endowed with anatomy more associated with pursuit, while the majority of the individuals possess anatomies more attributable to attractiveness. This is a curious behavior. It appears that a form of polymorphism might be emerging in these situations, whereby a genotype is easily expressed in one of two distinct phenotypes.
3.4.6. Beauty Should be in the Eye of The Beholder
For experimental purposes, the attractiveness criteria in these simulations were designed in a top-down, deterministic manner. To ground the simulation deeper, it would be better to allow beauty to be in the eye of the beholder and to be evolvable as well. This would require the design of pattern-recognition agents in creatures which size-up anatomies and motions of potential mates using their own genetically-determined criteria for beauty. These criteria should be sufficiently open-ended and variable to account for a very large space of possible criteria for beauty. Dawkins' concept of constrained embryology ideas come to mind again, only in this case, the method would be analytic instead of synthetic. Specific phenotypic features would be apprehended in a constrained way, through a "beauty filter."
next section:
Rendering
Rendering
